共济失调(Ataxia)是一类以运动协调性紊乱为主要特征的神经系统疾病,临床表现包括步态不稳、丧失平衡、吞咽困难、眼球运动异常、肌张力受损等。小脑功能障碍是导致共济失调发生的重要致病因素之一。
浦肯野细胞(Purkinje cell, PC)是小脑皮质中主要的GABA能神经元,也是从小脑皮质发出的唯一能够传出冲动的神经元。研究发现,小脑浦肯野神经元变性参与共济失调的发生发展过程。然而,参与共济失调的特异性投射环路及其潜在的病理机制仍不清楚。
2022年6月1日,南京医科大学基础医学院卢应梅教授课题组与药学院韩峰教授课题组合作在Signal Transduction and Targeted Therapy杂志在线发表最新研究论文“BOD1 regulates the cerebellar IV/V lobe-fastigial nucleus circuit associated with motor coordination”。此项研究首次报道了小脑IV/V叶到小脑顶核(Fastigial Nucleus,FN)投射环路异常是导致共济失调发生的重要病因之一,揭示了基于BOD1相关信号通路所导致的小脑IV/V叶浦肯野神经元至其下游投射的小脑顶核谷氨酸能神经元环路(IV/V lobePCs—FNCaMKIIα+)异常在共济失调发生发展过程中的分子机制。为探索预防共济失调等相关运动系统疾病的神经调控手段及临床治疗的分子药物靶点开发提供了强有力的依据。

小脑IV/V叶浦肯野细胞到FN CaMKIIα阳性神经元存在单突触投射
小脑顶核(Fastigial Nucleus,FN)可通过接受小脑皮质投射或发出神经纤维至大脑不同区域在协调运动中发挥关键作用。基于此,研究者借助病毒示踪技术和染料示踪技术明确小脑叶与FN之间的投射关系。
首先,作者将红色逆行示踪染料注射至WT小鼠FN,7天后发现,小脑IV/V叶到FN有投射通路(图1a)。随后,将顺行示踪病毒AAV-DIO-EGFP注射L7-Cre鼠(特异性在浦肯野细胞表达Cre)小脑IV/V叶,发现小脑IV/V叶浦肯野细胞(Purkinje cell, PC)到FN CaMKIIα阳性神经元之间的特异性投射(图1b-e)。进一步,作者借助逆行示踪病毒证实了IV/V lobePCs—FNCaMKIIα+投射通路(图1f-i)。
为表征IV/V叶到FN之间的突触连接,研究者将AAV-CaMKIIα-EGFP注射L7-Cre鼠FN,AAV-DIO-ChR2-mCherry注射IV/V叶。膜片钳记录显示,光遗传激活IV/V叶到FN的投射末端可导致FN CaMKIIα阳性神经元产生诱发性抑制性突触后电流(evoked inhibitory postsynaptic currents, eIPSCs),提示了小脑IV/V叶与FN CaMKIIα阳性神经元存在单突触投射(图1j-m)。
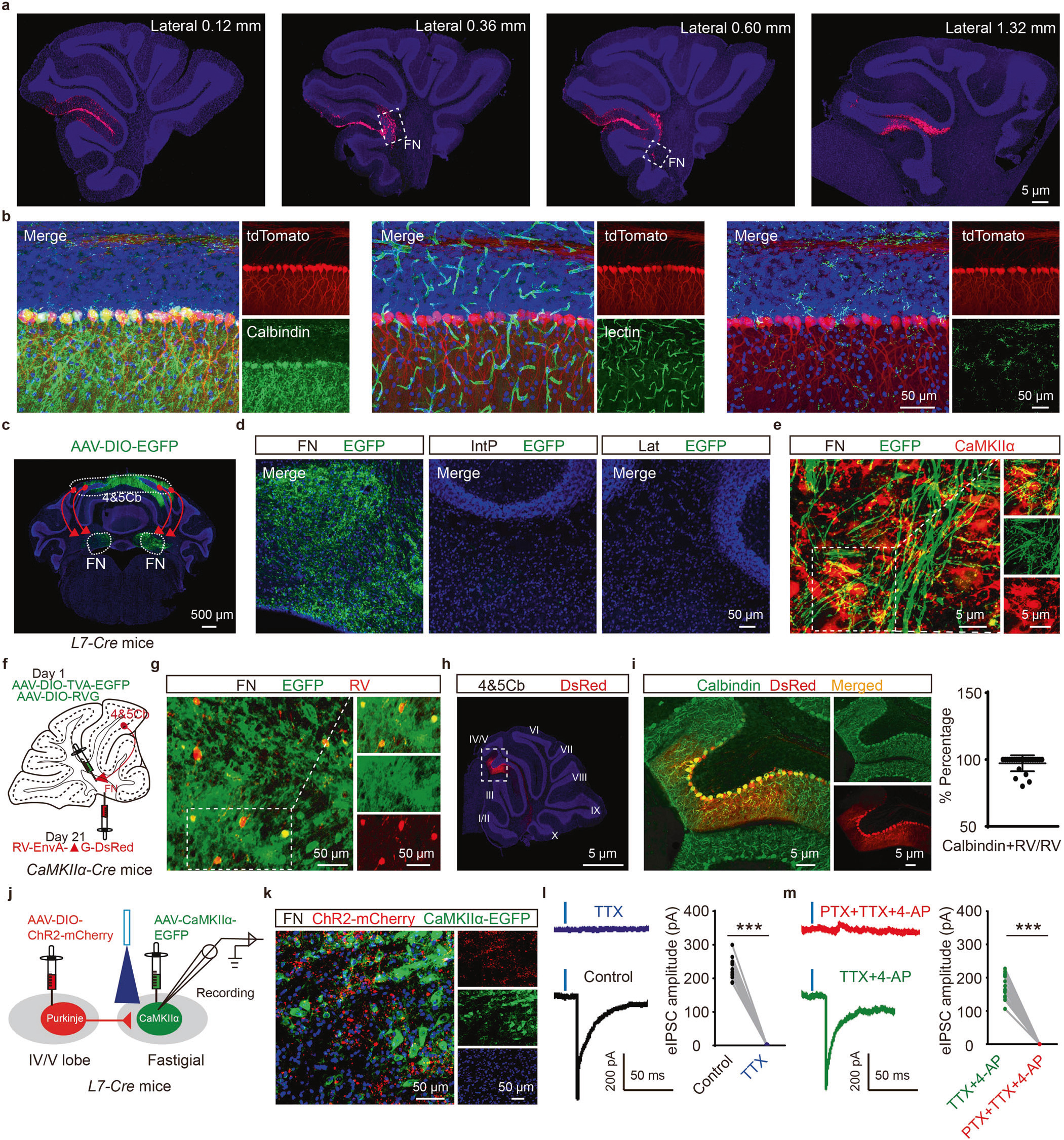
图1 IV/V lobePCs—FNCaMKIIα+投射通路
特异性抑制IV/V lobePCs—FNCaMKIIα+投射通路可导致共济失调样行为
为研究IV/V叶—FN环路在运动相关行为中的作用,研究者在L7-Cre鼠IV/V叶注射AAV-DIO-eNpHR3.0-mCherry病毒,对脑片进行全细胞膜片钳记录发现,光遗传抑制降低了IV/V叶浦肯野细胞的放电速率(图2a-e)。此外,多种行为学结果显示,光遗传抑制IV/V叶在FN的投射末端可诱发小鼠共济失调样行为(图2h-k)。同时,作者借助化学遗传学技术进一步验证了特异性抑制IV/V叶浦肯野细胞,可导致FN CaMKIIα阳性神经元过度激活,从而损伤小鼠运动协调能力,产生共济失调样行为。
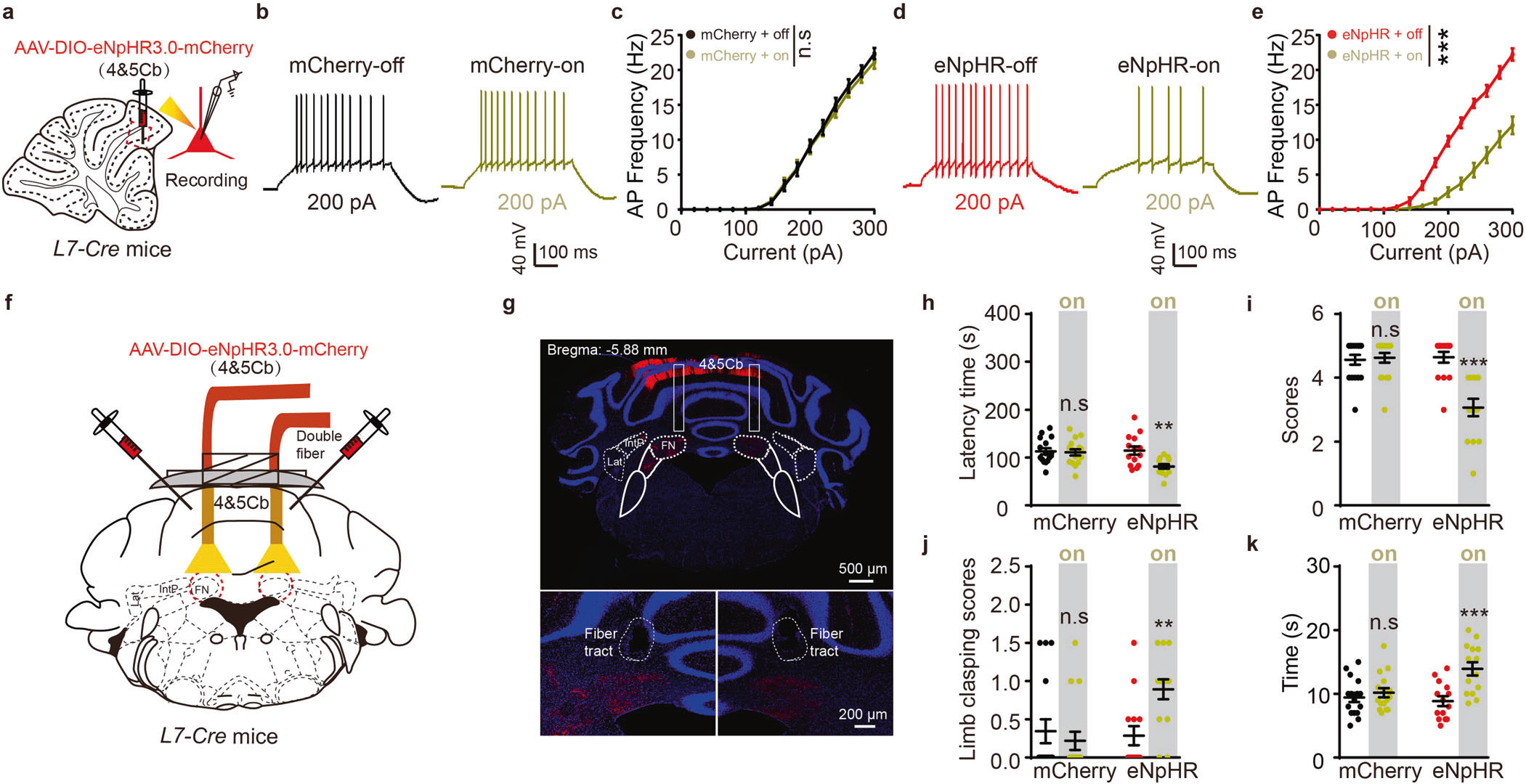
图2 光遗传抑制IV/V lobePCs—FNCaMKIIα+投射通路可导致共济失调样行为
为探讨IV/V lobePCs—FNCaMKIIα+投射功能异常的分子机制。研究人员通过转录组分析、GTEx(Genotype-Tissue Expression,基因型-组织表达)数据库和免疫荧光染色发现,BOD1在小脑浦肯野细胞中高表达(图3a-e)。
接下来,作者构建L7-Cre; BOD1f/f小鼠以确定特异性在浦肯野细胞中敲除BOD1基因对共济失调的影响。经多种行为学范式显示,L7-Cre; BOD1f/f小鼠出现共济失调样表型(图3f-m)。
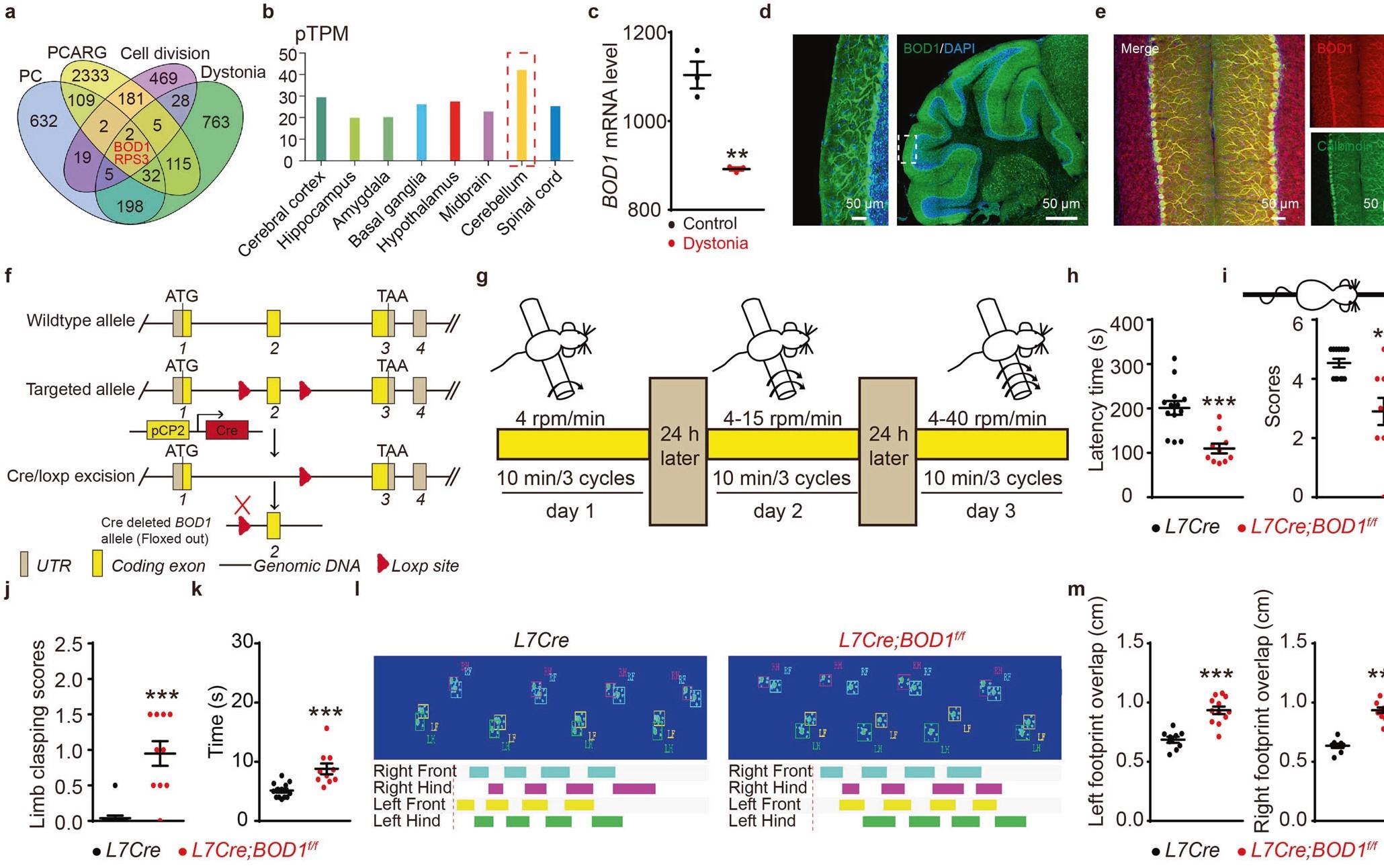
图3 特异性敲除浦肯野细胞中BOD1可导致共济失调
进一步,研究者借助AAV载体携带浦肯野细胞特异性启动子(AAV-L7-Cre-EGFP)条件性在小脑IV/V叶浦肯野细胞敲除BOD1,发现下调IV/V叶中BOD1的表达,可降低浦肯野细胞兴奋性,诱发小鼠表现出运动相关行为障碍(图4a-k)。
基于上述发现,研究人员分别在L7-Cre鼠和L7-Cre; BOD1f/f小鼠IV/V叶注射AAV-DIO-hM3Dq-mCherry病毒,借助化学遗传学技术激活浦肯野细胞,发现经CNO药物处理后,可改善由BOD1缺陷导致的小鼠共济失调样行为(图4l-p)。这些结果提示了,BOD1缺失影响小脑IV/V叶浦肯野细胞活性,导致IV/V lobePCs—FNCaMKIIα+投射功能异常,进而诱发小鼠共济失调样行为。
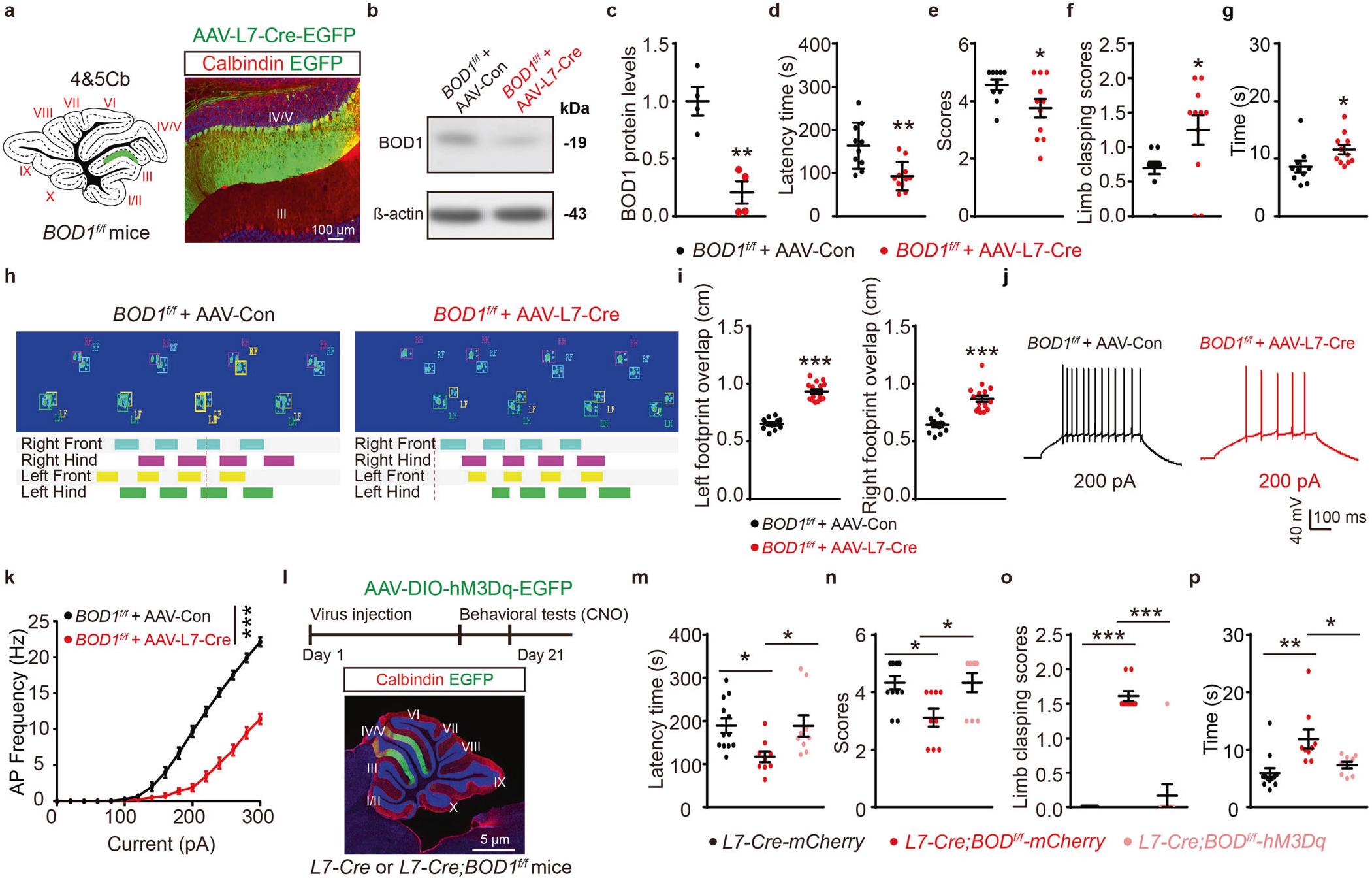
图4 BOD1调控IV/V lobePCs—FNCaMKIIα+投射环路
BOD1相关信号通路介导IV/V lobePCs—FNCaMKIIα+异常导致共济失调的分子机制
接下来,研究者借助稀疏标记病毒发现特异性敲除IV/V叶浦肯野神经元中BOD1可导致浦肯野细胞树突棘密度和树突分支明显降低。为研究其机制,作者分别在L7-Cre鼠和L7-Cre; BOD1f/f小鼠IV/V叶注射AAV-L7-EGFP,借助流式分选结合单细胞转录组分析发现,BOD1通过调节树突和树突棘相关基因影响浦肯野细胞树突形态发生改变(图5)。
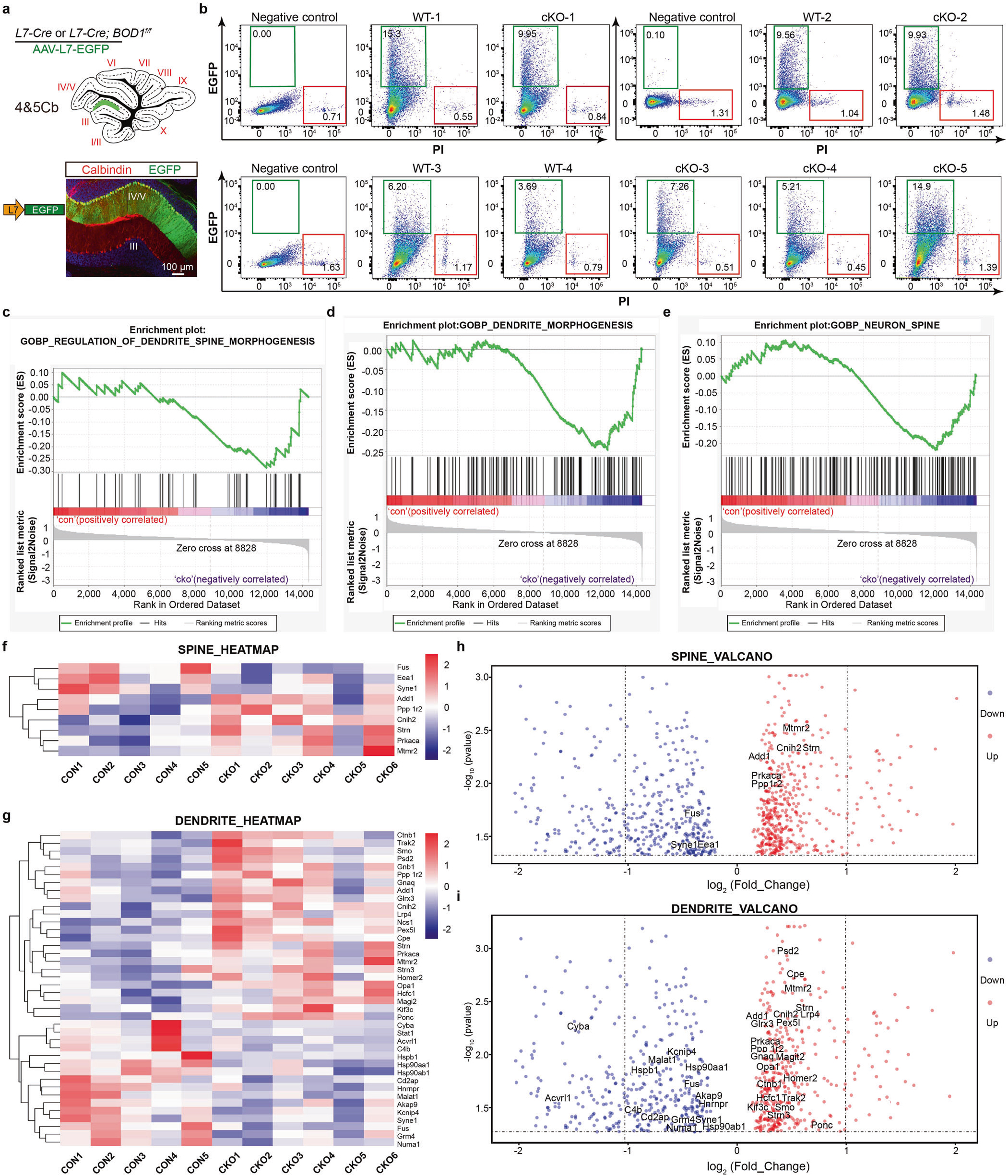
图5 BOD1通过调节树突和树突棘相关基因影响浦肯野细胞树突形态发生改变
最后,研究者将AAV-DIO-BOD1-mCherry注射于L7-Cre鼠和L7-Cre; BOD1f/f小鼠IV/V叶,发现上调BOD1的表达可增加IV/V叶浦肯野细胞兴奋性,逆转FN CaMKIIα阳性神经元异常放电现象,改善浦肯野细胞树突形态和共济失调样行为(图6)。
综上,IV/V叶浦肯野细胞中BOD1缺失通过调节树突和树突棘相关基因导致树突棘密度和树突分支降低,进而引起其活性下降,并通过影响IV/V lobePCs—FNCaMKIIα+投射环路,引起FN CaMKIIα阳性神经元异常放电,最终诱发共济失调发生。
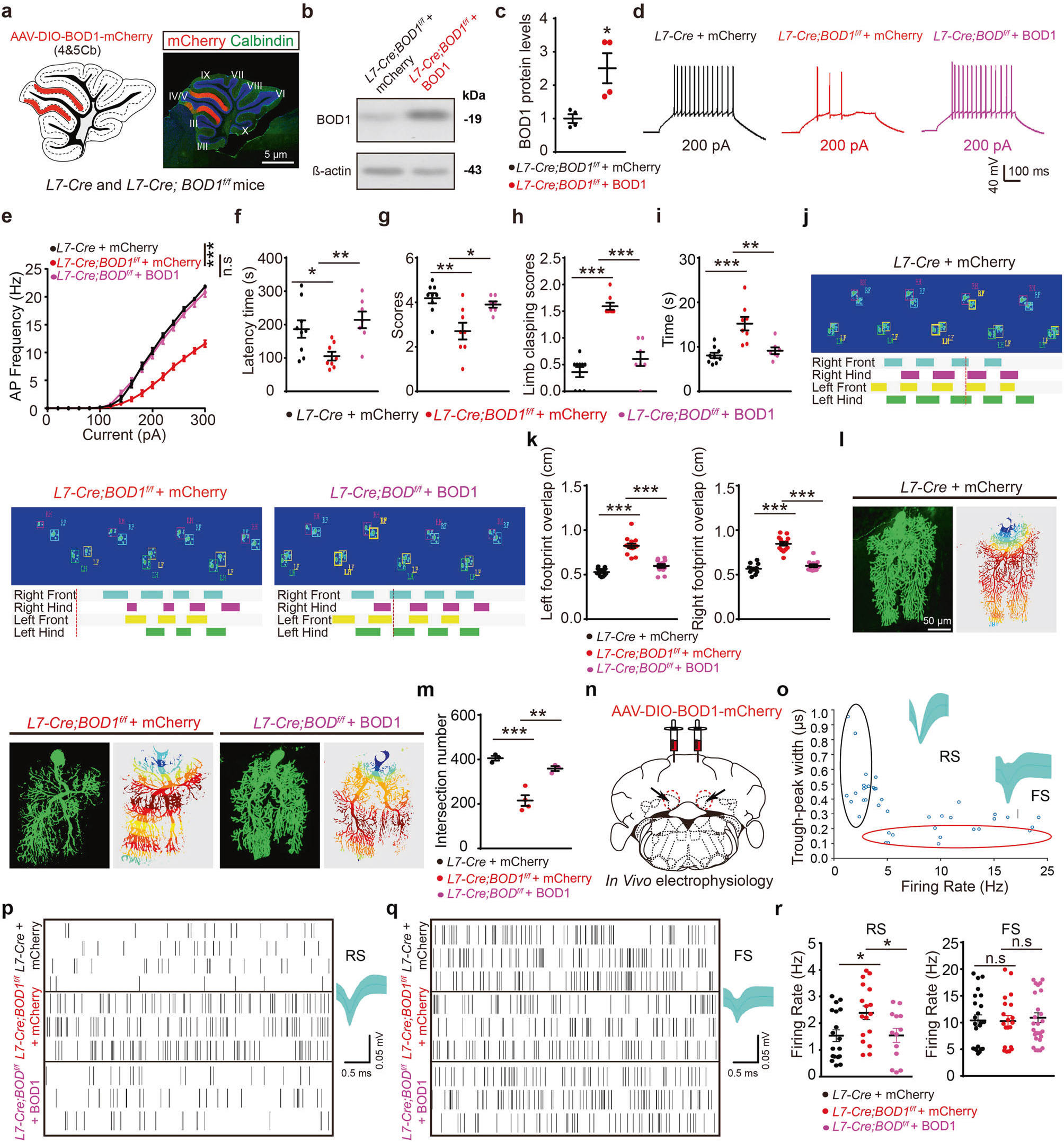
图6 上调IV/V叶浦肯野细胞中BOD1的表达可改善小鼠共济失调样行为
本文借助转基因小鼠、病毒示踪技术、膜片钳记录、光遗传学、化学遗传学、多种行为学范式、Cre-Loxp系统介导的条件性基因敲除、转录组分析等多种技术手段发现小脑IV/V叶浦肯野细胞中BOD1缺失可通过调节树突和树突棘相关基因导致树突棘密度和树突分支降低,进而引起其活性下降,并通过影响IV/V lobePCs—FNCaMKIIα+投射环路,引起FN CaMKIIα阳性神经元异常放电,最终诱发共济失调发生。此项研究找到了一条特异性介导共济失调发生的神经环路,为探索预防相关疾病的神经调控手段及临床治疗的分子药物靶点开发提供了强有力的依据。
南京医科大学药学院刘秀秀博士为该论文的第一作者,南京医科大学基础医学院卢应梅教授和药学院韩峰授为共同通讯作者。
和元生物有幸为研究者提供浦肯野细胞(L7)特异性启动子、光遗传、化学遗传学、Cre等AAV病毒载体,助力神经科学研究!